Grid Cells: The Brain's Coordinate System
In 2005, Hafting, Fyhn, Molden, and the Mosers discovered neurons in the rat medial entorhinal cortex that fire in a perfect hexagonal lattice as an animal moves through space — grid cells. This article explains how grid cells work, what their three key parameters (spacing, orientation, and phase) mean, how they implement path integration through self-motion signals, how they interact with hippocampal place cells, and why the hexagonal firing pattern may be a general representational format the brain uses to organize not just space but abstract knowledge.

June 14, 2026 · 8:10 AM
1 subscriptions · 19 items
The brain's internal GPS: how grid cells work
In 2005, a pair of neuroscientists at a Norwegian university had a problem they did not expect. They had implanted electrodes in the medial entorhinal cortex (MEC) of rats and let the animals explore large open arenas. As the recording data came in, the spike patterns looked strange — not the single compact hotspot you see from a hippocampal place cell, but a recurring burst that fired at multiple locations scattered across the floor. When Torkel Hafting plotted every position where a given cell fired during a thirty-minute session, the dots resolved into something none of them had seen before: a perfect hexagonal lattice, repeating like tiles across the entire environment. 1
The cell was firing at the vertices of equilateral triangles, each vertex equally spaced from its six neighbors, covering the floor from wall to wall. Hafting showed the map to his supervisors, May-Britt and Edvard Moser. The three of them stared at it. They had just found the brain's internal coordinate system.
Nine years later, May-Britt Moser, Edvard Moser, and John O'Keefe shared the Nobel Prize in Physiology or Medicine — O'Keefe for discovering place cells in 1971, the Mosers for discovering the grid cells that feed them. 2
What a grid cell actually does
A grid cell is a neuron in the medial entorhinal cortex (MEC) that fires whenever the animal occupies any vertex of a regular triangular grid spanning the full environment. 1 That is fundamentally different from a place cell. A place cell in the hippocampus fires at one location (or at most a handful of locations) per environment. A grid cell, by contrast, fires at dozens of locations — but those locations are not random. They form the most efficient tiling of a flat surface: the equilateral triangle lattice.

Each grid cell is characterized by three parameters:
- Spacing (or scale): the distance between adjacent firing fields, measured from vertex to vertex. This ranges from roughly 25 cm in the dorsalmost MEC to over 3 meters near the ventral tip — a tenfold range. 3
- Orientation: the angle at which the grid is rotated relative to the room. Neighboring cells at the same electrode share nearly the same orientation; cells recorded from different dorso-ventral locations can differ.
- Phase: where the grid vertices fall relative to some reference point. Two cells at the same electrode can have identical spacing and orientation yet tile different positions because their grids are offset from each other.
Critically, the spatial pattern persists in darkness and in the absence of visual landmarks — the light can be switched off and the hexagonal firing map barely flickers. 1 This was the first clue that grid cells are not reading the environment like a camera. They are computing position from within.
The autocorrelation signature
Neuroscientists confirm grid cell identity using an autocorrelogram: they take the firing-rate map and correlate it with itself at every possible spatial shift. A grid cell produces a distinctive hexagonally symmetric pattern — a hot central peak surrounded by six satellite peaks arranged at 60° intervals. This signature is unmistakable and has become the standard diagnostic.
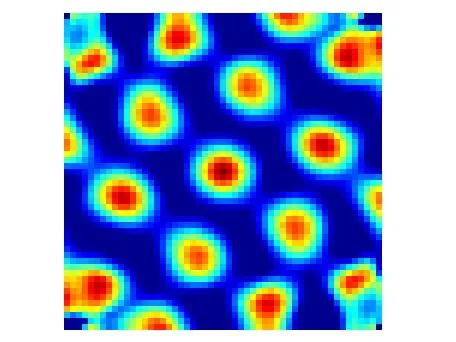
The autocorrelogram also provides an objective, continuous measure of "grid-ness," allowing researchers to score cells on a spectrum rather than forcing a binary yes/no classification. This tool has proved useful when searching for grid cells in animals other than rats — the hexagonal signature appears in mice, monkeys, and, most significantly, in humans navigating virtual environments. 4
How the grid is generated: path integration
The fact that grid patterns persist in darkness rules out a simple visual explanation. The leading account is path integration — the process of estimating current position by continuously integrating the speed and direction of self-movement. Imagine closing your eyes in a room: you can still keep rough track of where you are by counting steps and turns. Grid cells appear to implement a neural version of this dead-reckoning computation.
The MEC contains the raw ingredients. Layer II holds the densest population of pure grid cells. Deeper layers (III, V, VI) add conjunctive cells that carry both grid-pattern spatial information and head-direction tuning — they fire in a grid pattern only when the animal faces a particular compass direction. 5 Head-direction cells encode the animal's heading without any grid structure. Border cells fire when the animal runs along a wall. Together, these cell types in the MEC encode position, direction, and boundary information — exactly the quantities needed to calculate "where am I and where am I going."
Why a hexagonal lattice specifically? The equilateral-triangle tiling is the most efficient way to divide a plane into regions of equal area — it maximizes coverage per firing-field size and allows any two cells with offset phases to tile the environment without gaps or redundancy.

A prominent computational proposal, the continuous attractor network model, suggests that the hexagonal grid emerges when a sheet of recurrently connected MEC neurons tracks self-motion signals. In such models, the activity pattern on the neural sheet is maintained by the intrinsic dynamics of the network, shifted incrementally each time the animal moves. The hexagonal geometry arises because it is the most symmetric stable attractor solution for a two-dimensional sheet of neurons with a particular pattern of inhibitory and excitatory connections. 6
This model explains several observed facts: why the grid fires in darkness (the network sustains its pattern through internal dynamics), why all cells at a given dorso-ventral level share the same spacing (they belong to the same network module), and why cells at the same electrode have random phase offsets (their initial conditions differ).
The grid–place cell link
The anatomical relationship between the MEC and the hippocampus is tight and directional. Layer II of the MEC projects directly to the dentate gyrus and CA3 of the hippocampus via the perforant path. Layer III projects to CA1. The hippocampus, by contrast, projects back to the deeper layers of the entorhinal cortex. The circuit is not a passive relay but a feedback loop. 7
One of the most striking results from the Mosers' lab involved recording grid cells and place cells simultaneously while moving a rat between different environments. When the hippocampus showed complete remapping — its place cells reorganizing into a totally different map — the grid cells in the MEC did not remain static: their grids rotated and shifted unpredictably. But when the hippocampus showed only rate remapping (place fields staying put while firing rates changed), the grid cells showed no change at all. 8
This dissociation is telling: it implies that the MEC and hippocampus do not simply pass information in one direction. Both systems are capable of independent updates, and their joint state determines whether you are navigating a familiar or a new space.
A universal coordinate system?
The hypothesis that grid cells provide a universal metric for space — a kind of GPS chip in the brain — has been extended well beyond physical navigation. Several lines of evidence suggest the hexagonal firing structure reflects something more general.
In 2010, Doeller, Barry, and Burgess showed fMRI evidence of grid-like signals in the human entorhinal cortex during virtual navigation — the BOLD signal in the entorhinal cortex modulated at 60° periodicity in a way consistent with a hexagonal grid. 9 In 2013, Jacobs et al. recorded directly from epilepsy patients navigating a virtual town, finding grid-like activity in single neurons of the human entorhinal cortex. 4
More provocatively, Constantinescu, O'Reilly, and Behrens (2016) found hexagonal fMRI signals in the entorhinal cortex of humans navigating a conceptual space — a 2D space defined by the neck length and leg length of imaginary birds, with no physical movement at all. 10 If replicated, this would mean the hexagonal grid is not just a map of physical space but a general representational format the brain uses to organize any structured knowledge domain.
The DeepMind team published a parallel finding from artificial intelligence: when a recurrent neural network was trained to perform path integration — with no explicit instruction to develop grid-like activity — hexagonal units spontaneously emerged. Equipping artificial agents with these grid-like representations improved their performance on navigation tasks. 11 The brain may have discovered the same efficient representational solution that a gradient-descent optimizer converges on when given the same problem.
Open questions
Grid cell research has matured rapidly but several questions remain contested. The continuous attractor model is elegant, but the actual mechanisms generating the hexagonal pattern in biological tissue are still debated — particularly how the grid scale is set and whether theta-frequency oscillations are necessary (bats have grid cells without prominent theta oscillations 12). Whether the MEC is truly the site of path integration in humans (rather than a downstream reporter) remains unresolved, since entorhinal lesions in humans do not produce the same navigation deficits as in rodents. And the conceptual-space hypothesis — that grid cells organize abstract knowledge — is intriguing but requires more direct evidence from single-unit human recordings.
What is not in doubt is the broader architecture of the navigation system. O'Keefe's place cells give the hippocampus its location stamps. Moser's grid cells in the entorhinal cortex provide the metric — the sense of distance, direction, and displacement that makes one location distinguishable from every other. The two systems, anatomically adjacent and reciprocally connected, together build the internal map that makes navigation possible.
Landmark paper: Hafting T, Fyhn M, Molden S, Moser M-B, Moser EI. "Microstructure of a spatial map in the entorhinal cortex." Nature 436:801–806 (2005). DOI: 10.1038/nature03721. PMID: 15965463. 1
Course connection: MIT 9.13 The Human Brain (Nancy Kanwisher) — Navigation and Spatial Memory module. Related lectures at nancysbraintalks.mit.edu.
References
- 1Hafting et al. 2005, Nature
- 2Nobel Prize 2014 announcement
- 3Brun et al. 2008, Science
- 4Jacobs et al. 2013, Nature Neuroscience
- 5Sargolini et al. 2006, Science
- 6Burak & Fiete 2009, PLOS Comp. Biol.
- 7Van Strien, Cappaert & Witter 2009, Nature Reviews Neuroscience
- 8Fyhn et al. 2007, Nature
- 9Doeller, Barry & Burgess 2010, Nature
- 10Constantinescu, O'Reilly & Behrens 2016, Science
- 11Banino et al. 2018, Nature
- 12Yartsev, Witter & Ulanovsky 2011, Nature
Add more perspectives or context around this Post.